Introduction
Two panicked thoughts crossed my mind after I heard the news about Ornithoscelida on March 22. The first was that Derek was going to beat me to the punch in writing a post about the phylogenetic revision of dinosaurs. The second was that I was going to have to rewrite a lot of my current research. I was right about one of those worries (if delightedly so) [1]. As for the other: well, let's try to make some lemonade out of these lemons.
The research in question is intended to break down the traditional distinction between body fossils and trace fossils [2]. Dinosaurs have been my go-to example for how paleontologists draw different inferences from different kinds of fossil, but the new phylogenetic revision--the "Ornithoscelida hypothesis"--undermines that example. Re-examining some assumptions might yield a more helpful distinction than the traditional one. Let's give it a shot!
Retreading the trackway
Here's a summary of what I've found since first writing about this topic last year:
- Paleontologists maintain separate taxonomies for body fossil and for trace fossils.
- Following the International Code of Zoological Nomenclature, vertebrate paleontologists give priority to body fossil taxon names over trace fossil taxon names. This is because of difficulties in connecting trace species to tracemaker species. The names of body fossil taxa have equal priority with the names of extant taxon names.
- Following the International Code of Nomenclature for Algae, Fungi, and Plants, paleobotanists give equal priority to all fossil taxon names, which is lower than the priority given to extant taxon names. This is because of difficulties in connecting all of the relevant organisms' life stages to a single species.
- It turns out that the ICZN's distinction is vague and, to be frank, often nonsensical. To wit: dinosaur feather imprints count as body fossils--the former type specimen of Archaeopteryx lithographica is one such example--but skin imprints count as trace fossils.
- Like paleobotanists, vertebrate paleontologists often have difficulty recognizing different life stages of a single body fossil species. Want all the vitriol of a political debate without any of the practical consequences? Try following the "Nanontyrannus" hashtag on Twitter.
My goal has been to argue against the vertebrate paleontologists' conventional prioritization of body fossils over trace fossils. One significant obstacle to overcome has been the discrepancy between phylogenies for dinosaurs and their traces.
Consider the chart below, which shows the major groups of dinosaur ichnotaxa:
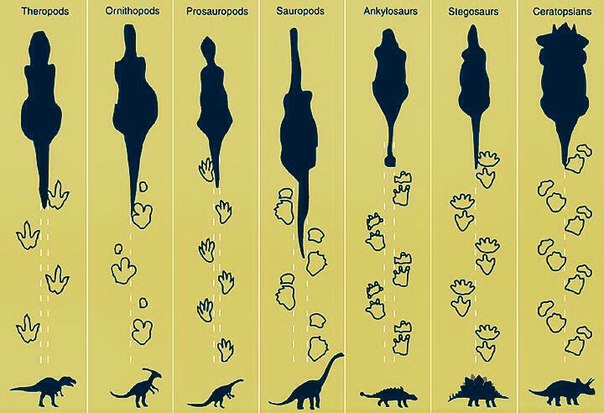
Image from the University of Texas at El Paso.
Comparing similarities between these tracks, the most easily quantified grouping (among three-toed tracks) would place the theropods with ornithopods and their relatives, excluding sauropods in a separate group. But grouping theropods with ornithopods was supposed to be inconsistent with the "real" dinosaur family tree, pictured below:

Image from the Tetrapod Zoology blog.
The supposedly true evolutionary history of dinosaurs groups theropods with sauropods, excluding ornithopods and their relatives (i.e., the Ornithischia). If this is the dinosaur family tree that evolution really gave us, and if the family tree formed by comparing dinosaur tracks is significantly different, then shouldn't that just be too bad for the tracks? In other words, body fossil taxonomy--which formed the basis of the "real" tree--clearly has priority over trace fossil taxonomy.
Clearly. Until March 22.
Ornithoscelida steps up
Thomas Henry Huxley originally suggested the name "Ornithoscelida" for what we now call the Dinosauria, citing the avian characteristics common to all members of the group [3]. One reason that his terminology fell out of a favor was opposition to his inclusion of birds within the group and, consequently, within Reptilia. In any event, the name didn't stick until just a few weeks ago, when Baron, et al. (2017) resurrected it in their phylogenetic revision of dinosaurs.

Image from the Tetrapod Zoology blog.
This revised phylogeny classifies theropods with ornithopods and their relatives in a new group named Ornithoscelida, excluding sauropods (as well as some dinosaurs formerly classified as theropods). Where have we heard that before?
Baron, et al's "Ornithoscelida" is not the same as the group that Huxley originally designated with that name, but it does correlate with the grouping formed by dinosaur trace fossils. To be fair, Baron, et al. don't mention tracks at all in their analysis. Their conclusions follow from analysis of 457 traits across 74 dinosaur body fossil taxa. This analysis may or may not withstand further scrutiny--apparently the addition of other traits or taxa can restore the traditional view--but it does show that the family tree derived from analysis of dinosaur tracks is not an unreasonable one per se.
One might argue that it would be unreasonable to draw inferences about evolutionary history from fossils of tracks, which offer relatively low-resolution taxonomic information. But it's worth noting that Baron, et al did draw inferences from integumentary impressions--one of the variety of traits that may unite theropods and ornithischians--which count as body fossils. Why is it more reasonable to draw inferences from impressions of one part of an organism's body than it is to draw inferences from impressions of another part of the organism's body? Sure: some impressions are more informative than others, but impressions themselves--traces--can be informative enough.
A step in the right direction

The world's largest known dinosaur track (top), provisionally attributed to the mellifluously-named taxon "Broome Sauropod Morphotype A." From Salisbury, et al (2016).
Nevertheless, vertebrate paleontologists tend to maintain that trace fossils 'contribute very little to the study of dinosaur phylogeny.' Feather impressions do make such contributions because they happen to fall on the "right" side of the body fossil/trace fossil divide. This strikes me as capricious.
How can we do better? The answer occurred to me earlier this week, when a student asked me to define the term "phenotype." I gave a standard answer--the physical manifestation of an organism's genetic information--which prompted discussion about phenotypic plasticity. After all, the organism's genotype alone isn't sufficient for it's phenotype; the latter is the product of the former's interaction with the environment. "Product of a genotype's interaction with the environment" also pretty accurately summarizes the causal history of a feather impression. Here's the kicker: so too does it describe the causal history of a footprint.
The line between body fossils and trace fossils is traditionally drawn where the organism's body supposedly ends, but don't let your sense of proprioperception fool you: that boundary is always a vague one. The boundary at which an organism's phenotype ends is similarly fuzzy--some of you may recall that Richard Dawkins had some controversial things to say on the matter--but it's a more useful one for drawing phylogenetic inferences. Footprints encode phylogenetic information at a very low resolution, but that low-resolution information can be incredibly helpful when you're drawing up a large-scale revision of dinosaurs' evolutionary history.

A footprint from New Mexico preserving the 'unequivocal and diagnostic theropod hallux.' Lockley, et al (2011) argue that 'given the size, the attribution to genus Tyrannosaurus is convincing and as this is arguably a monospecific genus, it is likely the track of a T. rex.' The print is still classified in the ichnotaxon Tyrannosauripus pillmorei. Image courtesy of Wikipedia Commons.
Some fossils classified as traces, such as tracks or skin impressions, preserve part of an organism's phenotype. Other trace fossils, such as coprolites or burrows, are less obviously phenotypic manifestations (see the link given above for more discussion of Dawkins' "extended phenotype" concept). The former kinds of trace fossil should be phylogenetically informative where the latter might not be; in any event, it's clearly false to say that no trace fossils have much to contribute to phylogenetic reconstruction.
I propose, then, to get rid of the trace fossil/body fossil distinction altogether. It's at least as arbitrary as the distinction between body and environment (perhaps the one has something to do with the other?). Replace it with a distinction between phenotype-related fossils and others. Feet, feathers, and skin are one thing; poop and bedrooms are another. This new distinction preserves the motivation behind the old one--presumably, to recognize when a paleontologist is studying an organism and when the paleontologist is studying something else--but it also captures more of theoretical interest while avoiding strange inconsistencies.
Bonus: a pheno-fossil/non-pheno-fossil distinction gives us one more datapoint in the debate over Ornithoscelida, which is certainly shaping up to be a boon for philosophy of paleontology. Don't say that we didn't try to get you in on the ground floor!
Notes
[1] If you haven't yet read Derek's typically excellent essay from last week, then I strongly encourage you to do so now. I hope he doesn't mind my paying homage to his work with the title of my follow-up.
[2] Derek beat me to the punch on that one, too, in his 2007 book. To be fair: I was in grad school at the time.
[3] Hence the term "ornithoscelidaphobia," which means "fear of dinosaurs." (The term also gives an incredible branding opportunity for Baron, et al, who now have a ready-made phrase to describe their intellectual opponents.)
Works cited
- Baron, M.G., Norman, D.B., and Barrett, P.M. (2017). A new hypothesis of dinosaur relationships and early dinosaur evolution. Nature, 543: 501-512.
- Lockley, M., Janke, P. R., & Triebold, M. (2011). Tracking Tyrannosaurus: notes on purported T. rex tracks. Ichnos, 18: 172-175.
- Pemberton, G. and Frey, R.W. (1982). Trace Fossil Nomenclature and the Planolites-Palaeophycus Dilemma. Journal of Paleontology, 56: 843-881.
- Salisbury, S. W., Romilio, A., Herne, M. C., Tucker, R. T., & Nair, J. P. (2016). The Dinosaurian Ichnofauna of the Lower Cretaceous (Valanginian–Barremian) Broome Sandstone of the Walmadany Area (James Price Point), Dampier Peninsula, Western Australia. Journal of Vertebrate Paleontology, 36, 1-152.
- Turner, D. (2007). Making Prehistory: Historical Science and the Scientific Realism Debate. New York, NY: Cambridge University Press.